【トピックス】
オルガネラ膜融合:試験管内完全再構成による新展開
三間穣治
阪大・蛋白研
1.はじめに
出芽酵母Saccharomyces cerevisiaeからヒトをはじめとする高等動物に至るまで、全ての真核細胞は、細胞周期や細胞外環境などの生育状態に応じて、個々の細胞内オルガネラ膜の形態をダイナミックに変化させながら生命活動を行っている。本稿の主題である「オルガネラ膜融合」は、これらの「オルガネラ膜ダイナミクス」において必須かつ根源的な生体反応であり、個々のオルガネラの生命機能とも密接に関係している。真核細胞の「オルガネラ膜融合」研究はこれまで主に、1970年代よりNovickとSchekmanらによる出芽酵母温度感受性“SEC”変異株スクリーニングなどの遺伝学的解析、またはBalchとRothmanらによる哺乳動物ゴルジ体間膜輸送アッセイに代表される、単離オルガネラを用いた生化学的解析によって大きく進展した1)。そしてこれらの研究手法によって、「オルガネラ膜融合」に関与するタンパク質因子として、NSF (N-ethylmaleimide sensitive factor)、α-SNAP (Soluble NSF association protein)、SNARE (SNAP receptor)、SM (Sec1/Munc18) ファミリータンパク質などが同定されてきた1)。近年ではさらに、これら上記のタンパク質因子群に加えて、ホスホイノシチド (イノシトールリン脂質)、ステロール類、ジアシルグリセロール (DAG)、ホスファチジン酸 (PA) など、特異的な脂質群がオルガネラ膜融合に重要であることも示され、それらの役割も明らかになりつつある2)。本稿では、Wickner (米国ダートマス大学) らが「オルガネラ膜融合」の研究モデルとして用いて、多くの成果を挙げた、「出芽酵母液胞オルガネラ膜融合」を中心に概説する3)。そして特にWicknerと筆者らが最近成功に至った、「精製タンパク質と人工脂質二重膜リポソーム」を用いた化学的に純粋な実験系である、「液胞オルガネラ融合の試験管内完全再構成系」に焦点を当て、その最新の成果を紹介する4)。
2.SNAREの発見、SNARE仮説、そして酵母液胞オルガネラ膜融合
前述のようにRothmanらは1980年代後半から90年代前半にかけて、動物細胞より単離したゴルジ複合体から試験管内ゴルジ層間輸送アッセイ法を確立し駆使することで、膜融合に関与するタンパク質因子を次々と同定した1)。まずNSFを細胞質画分から同定し、次にNSFのアダプタータンパク質としてα-SNAPを、そしてNSF/α-SNAPのオルガネラ膜上のレセプター分子として、SNAPレセプター、つまりSNAREを発見するに到った5)。SNAREはその後、動物のみならず、植物、そして酵母を含め、全ての真核生物において保存される、細胞内膜融合に必須のタンパク質ファミリーであることが、生化学、遺伝学、細胞生物学的手法によって示されてきた5)。その中で、1998年にはRothmanらが、SNAREの精製タンパク質と人工脂質二重膜リポソームから、SNARE依存性膜融合を試験管内完全再構成することに初めて成功した6)。そこでは、シナプス神経伝達に必須のSNAREタンパク質と、ホスファチジルコリン (PC) ・ホスファチジルセリン (PS) のみから構成されるシンプルな脂質組成で、SNAREプロテオリポソームを再構成した。そしてこれらのSNAREプロテオリポソームが、生理的温度条件下で、プロテオリポソーム間でトランスSNARE複合体を形成、自発的にドッキング、そして膜融合を起こすことを報告した6)。この再構成SNAREプロテオリポソーム融合の結果が、実験的な基盤となり、「SNAREタンパク質が、膜融合に必須かつ十分で、膜融合の最小マシナリーである」という、いわゆる「SNARE仮説」が提唱され、現在も広く受け入れられている。
しかしその一方でWicknerらは、1990年代より酵母液胞 (動物細胞のリソソームに相当するオルガネラ) をモデルとしたオルガネラ膜融合研究を進める中、Rothmanらの「SNARE仮説」とは異なり、SNAREだけでは膜融合に十分でなく、他の多様な膜融合因子群も重要であることを示してきた3)(図1)。実際には、出芽酵母細胞より単離精製したオルガネラ、液胞を材料に、オルガネラ膜融合の生化学的アッセイ法を確立3)、各候補因子に対する特異的阻害剤を添加するなど、様々な反応条件下でこの膜融合アッセイを駆使することで、液胞融合の必須因子を探索してきた。同定された必須タンパク質因子の中には、液胞に局在する3つのQ-SNAREタンパク質 (Vam3p、Vti1p、Vam7p) と1つのR-SNAREタンパク質 (Nyv1p) に加えて、SNAREシャペロンとしてSec17p (α-SNAPの酵母ホモログ)、Sec18p (NSFの酵母ホモログ)、HOPS複合体 (Homotypic fusion and vacuole protein sorting complex、SMファミリータンパク質Vps33pをサブユニットの1つとして含む)、そしてRab GTPアーゼYpt7pを含んでいる (図1)。さらには脂質因子としては、ステロール類のエルゴステロール (ERG)、DAG、ホスホイノシチドとして、ホスファチジルイノシトール3リン酸 (PI(3)P) とホスファチジルイノシトール4,5ビスリン酸 (PI(4,5)P2) が同定され、様々な脂質もオルガネラ膜融合に関与することが示されてきた3)。これら単離したオルガネラを用いた生化学的解析の他にも、遺伝学・細胞生物学的解析として、酵母1遺伝子破壊ライブラリーを利用した「液胞形態変化」のin vivo網羅的スクリーニングでは、同定された因子の欠損酵母の多くで、膜融合不全の表現型とされる液胞断片化 (フラグメンテーション) が観察されている7)。つまりは、酵母液胞をモデルとしたオルガネラ膜融合の一連の研究では、SNAREのみならず、そのシャペロンや特異的脂質分子など多種多様な因子群が協同的に膜融合を引き起こしている、という結論が導かれてきた。
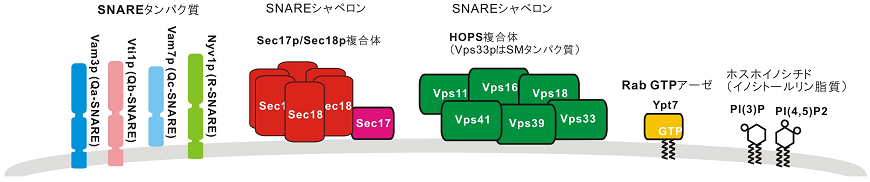
図1 出芽酵母液胞オルガネラ膜融合のタンパク質因子・脂質因子
3.酵母液胞オルガネラ膜融合の試験管内完全再構成
Rothmanらによる前述の「再構成SNAREプロテオリポソーム膜融合」は、精製SNAREタンパク質と合成脂質のみから構成される魅力ある実験手法であったが、そこから得られた「SNAREが膜融合に必須かつ十分である」とするSNARE仮説は、Wicknerらの酵母液胞をモデルにした「単離オルガネラの生化学アッセイ」や「オルガネラ形態のin vivo解析」で得られた実験結果・知見とは大きなギャップが存在していた。そこで、Wicknerと筆者らは、Rothmanらによる従来のSNAREのみから再構成されたプロテオリポソーム膜融合ではなく、実際の細胞内の生理的環境をより再現した、SNAREシャペロン群とATPを含めた、新たなプロテオリポソーム再構成系の構築に挑戦した4)。それにより、これまで用いられてきた非生理的で単純な脂質組成 (PC (85%) /PS (15%)) では、SNAREシャペロン/ATP存在下でも、液胞SNAREプロテオリポソームの膜融合は起こらないことを示した一方で (図2)、実際のオルガネラ酵母液胞に近い生理的脂質組成 (PC (45%) /ホスファチジルエタノールアミン(PE) (18%) /ホスファチジルイノシトール(PI) (18%) /PS (4%) /PA (2%) /ERG (8%) /カルジオリピン(CL) (2%) /DAG (1%) /PI(3)P (1%) /PI(4,5)P2 (1%)) に変えることによって、SNARE、SNAREシャペロン (Sec17p/Sec18p/HOPS)、そしてATP、これら全ての膜融合因子に依存する、効率的で速いプロテオソーム膜融合を得ることができた (図2)。このことは、SNAREは膜融合の最小マシナリーではなく、SNAREに加えてSNAREシャペロン群/ATPも膜融合のコアマシナリーであることを支持し、さらには生理的脂質組成が、これら膜融合タンパク質因子群の機能発現に重要な役割を持つことを明らかにしている4)。
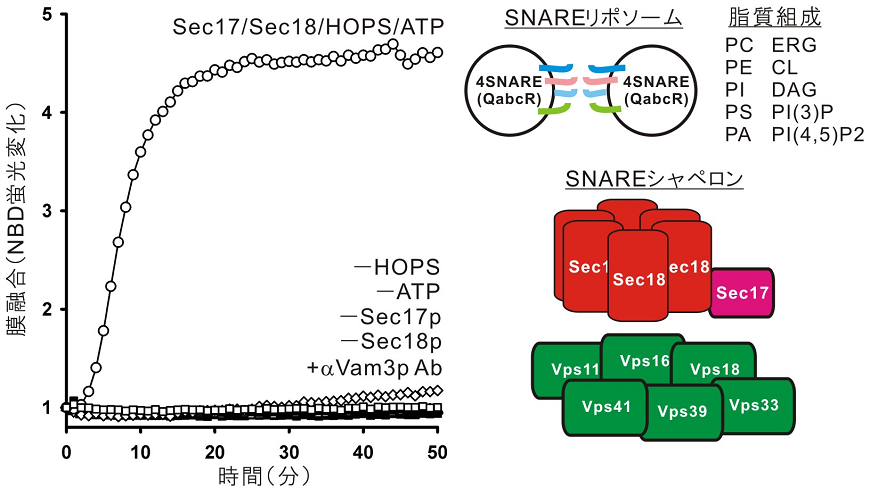
![]()
この「液胞オルガネラ膜融合」の「試験管内完全再構成系」を使い、次にWicknerと筆者は、必須脂質因子の1つ、ホスホイノシチドの「膜融合」における特異的な役割を明らかにすることを目指した8,9)。脂質組成を自由自在に操作できる、という再構成プロテオリポソーム系ならではの特徴を活用し、ホスホイノシチドを含めて脂質組成が異なる様々なSNARE再構成プロテオリポソームを調製、それらの膜融合活性を網羅的に測定した。そしてこれまで、「液胞SNARE/SNAREシャペロン (Sec17p/Sec18p/HOPS) 依存性膜融合において、PI(3)Pが、最も顕著に膜融合速度を促進する特異的なホスホイノシチドであり」(図3A)、さらに「PI(3)Pの機能 (SNARE/SNAREシャペロンの活性化) は、生理的な液胞バルク脂質群によって最適化されること」(図3B)、これら2点が新たに見出された8,9)。ただし、PI(3)Pをはじめホスホイノシチドが具体的に、オルガネラ膜上でどのようにSNARE/SNAREシャペロンといった膜タンパク質因子群に作用するかなど、詳細な分子機構については依然全くの不明であり、それらへの実験的介入は今後の大きな研究課題の一つとなるであろう。
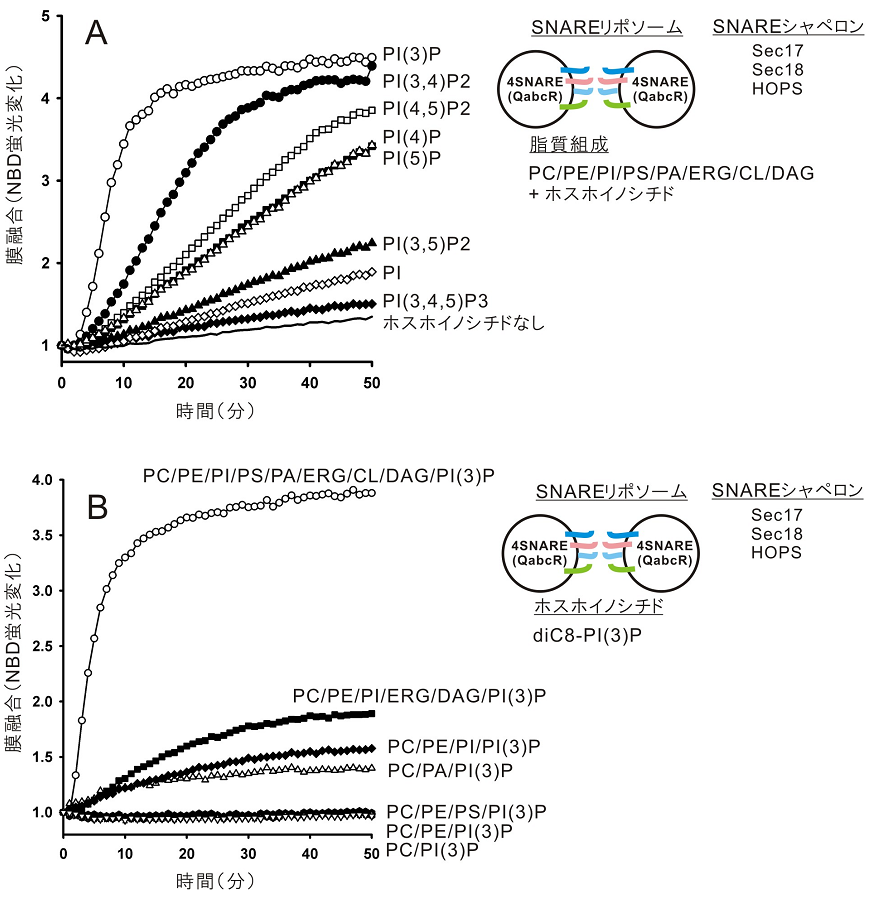

4.おわりに
今回紹介した、SNARE、SNAREシャペロン、生理的で複雑な脂質組成、そしてATPを含む「酵母液胞オルガネラ膜融合の試験管内完全再構成」は、多種類の精製膜タンパク質の調製、機能的なプロテオリポソームへの再構成、そして定量的な機能アッセイ法の確立と、技術的な面で非常に困難で挑戦的であったが、酵母液胞融合だけでなく他のオルガネラ膜融合・膜動態の研究にも波及する多くの新しい概念を生み出すことが出来た。そして「試験管内完全再構成系」の有用性は現在進行形で発展を続けており、例えば、Wicknerらはその後、液胞Rab GTPase Ypt7pを加えた新しい液胞融合再構成系を発表10,11)、そしてZerialらは、Rab5 GTPase、Rab5エフェクター群、そしてエンドソームSNAREを含むエンドソーム膜融合の試験管内完全再構成に成功している12)。今後は、さらに生理的なオルガネラに近づく進化した試験管内完全再構成系が構築され、「膜融合」のみならず、「膜出芽・分裂」、「膜変形」、「膜移動」も含めて、オルガネラ膜ダイナミクスの統合的な理解に向かう大きなブレイクスルーが生まれる事を期待している。
文献
1) Bonifacino, J. S., Glick, B. S.: Cell, 116, 153 (2004).
2) Wickner, W., Schekman, R.: Nat. Struct. Mol. Biol., 15, 658 (2008).
3) Ostrowicz, C. W., Meiringer, C. T. A., Ungermann, C.: Autophagy, 4, 1 (2008).
4) Mima, J., Hickey, C. M., Xu, H., Jun, Y., Wickner, W.: EMBO J., 27, 2031 (2008).
5) Jahn, R., Scheller, R. H.: Nat. Rev. Mol. Cell Biol., 7, 631 (2006).
6) Weber, T., Zemelman, B. V., McNew, J. A., Westermann, B., Gmachl, M., Parlati, F., Söllner, T. H., Rothman, J. E.: Cell, 92, 759 (1998).
7) Seeley, E. S., Kato, M., Margolis, N., Wickner, W., Eitzen, G.: Mol. Biol. Cell, 13, 782 (2002).
8) Mima, J., Wickner, W.: Proc. Natl. Acad. Sci. USA, 106, 16191 (2009).
9) Mima, J., Wickner, W.: J. Biol. Chem., 284, 27114 (2009).
10) Hickey, C. M., Stroupe, C., Wickner, W.: J. Biol. Chem., 284, 16118 (2009).
11) Stroupe, C., Hickey, C. M., Mima, J., Burfeind, A. S., Wickner, W.: Proc. Natl. Acad. Sci. USA, 106, 17626 (2009).
12) Ohya, T., Miaczynska, M., Coskun, U., Lommer, B., Runge, A., Drechsel, D., Kalaidzidis, Y., Zerial, M.: Nature, 459, 1091 (2009).
![]()