【トピックス】
微細藻類ユーグレナのバイオ燃料生産の可能性
鈴木健吾、石川孝博
株式会社ユーグレナ、島根大
1.はじめに
将来における食料やエネルギーの枯渇が予想される現在、地球上の物質循環を実現させることを想定して藻類バイオマスの食料や燃料などへの利用が注目されている。日本においては、数ある微細藻類の中でも細胞内に特殊な代謝機構を有し、動物性タンパク質や多種のビタミンを生産するユーグレナに1970年代から注目が集まってきた。ユーグレナは、日本では「ミドリムシ」の和名で広く知られ、一般に沼、池、水田などの淡水に生息している水中を浮遊する微細藻類の一種である (図1)。ユーグレナは葉緑体を有し光合成を行う植物性の性質を有しながら、鞭毛や細胞収縮による運動性をもつなどの動物的な特徴をも示すことから、古典的な分類学上において植物と動物の両方の分類に属する特殊な微生物として知られる1)。数十以上存在するユーグレナ属の中でも、特にEuglena gracilisは、溶存二酸化炭素濃度が高く低pHの環境条件に対して適応可能であることから他生物の混入を防ぎつつ培養することが可能であり2)、また、β-1,3-グルカンを合成しパラミロンと呼ばれる顆粒として蓄えること3)や、特殊な条件下でワックスエステルを合成する4)など特徴的な代謝経路を有しており、産業上それらが活用できる可能性が高いと考えられてきた。

図1 ユーグレナの光学顕微鏡画像
著者の一人が所属する株式会社ユーグレナでは長年期待されていたユーグレナの大量培養を実現し、2007年には産業利用も本格的に開始している。水中で培養したユーグレナは遠心分離機などを用いて濃縮、乾燥させて粉末状にした後に、主に食品として加工している。ユーグレナは食品としては、幅広い栄養素を有しており、消化・吸収を妨げる細胞壁がないことから消化率も高い。またユーグレナ特有の成分であるパラミロンは食物繊維様の働きや抗腫瘍活性および免疫機能の向上など様々な機能性を有することが明らかになっており、ユーグレナの食材としての価値を高めている5)。本稿では、さらなるユーグレナの産業利用の試みについて紹介する。
2.ユーグレナによる排気ガス中CO2削減の可能性について
藻類を利用したバイオマス生産の大きな利点の一つは、直接水中に溶け込ませた二酸化炭素を炭素源として利用することが可能である点である。前述したようにユーグレナは他の藻類より高濃度の二酸化炭素を溶解した培養液中でも増殖可能であることから2)、ユーグレナの産業利用においては、ユーグレナの培養による二酸化炭素固定と食料生産を同時に達成することが一つの大きな目標となっている。代表的な例として、1990年代に近藤次郎らによる「K プロジェクト」においてユーグレナの大量培養を目指した構想が提示されている6)。このプロジェクトでは、発電所で発生する二酸化炭素を利用し、発光ダイオード (LED) を用いて太陽光の不足を補うことによりユーグレナの光合成を促進することで、二酸化炭素固定と同時に食料を生産できると期待された。
Kプロジェクトを現実のものとするため、筆者らは沖縄電力の金武火力発電所にユーグレナの培養槽 (容量500リットル) を設置し、発電所からの排気ガスをユーグレナ培養槽に通気する実証試験を継続して行ってきた7) (図2上)。ボイラーで石炭を燃焼することにより発生した排気ガスは、脱硝装置、電気集塵器、脱硫装置を経ることにより、硫黄酸化物、窒素酸化物、煤塵等が排出基準値以下に抑えられる。排気ガスに含まれる二酸化炭素濃度は10〜15% (通常の大気中二酸化炭素濃度は0.04%) であるが、それらの浄化処理工程を経た後の排気ガスを引き込み培養したところ、実際に空気だけを通気した場合よりも良好な増殖がみられる結果を得た (図2下)。また、藻体の回収と培地の入れ替えを行うことで、半連続培養を行うことが可能であることも確認した。上記の実証試験では受光面積が増殖の限界を決定しているため、現在得られている細胞濃度は実験室内で得られる値の10分の1程度であり、この改善が実用化に向けての鍵となる。そのために受光面積を拡大できるような培養のモジュールの接続を検討し住友共同電力とさらなる共同研究を行っている。現段階で培地1 mlあたりユーグレナ5 × 105個程度の細胞濃度まで増殖させることに成功した。このような排気ガスを利用した高密度培養では密閉空間にて培養を行うことが肝要であり、外気と接しない培養槽の構築を実現すれば、外来遺伝子を導入した組み換え体ユーグレナも拡散をさせることなく培養できる可能性が期待できる。
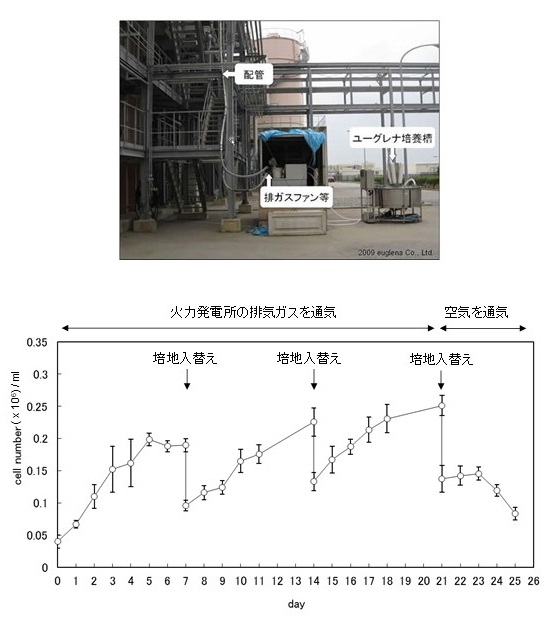
図2 石炭火力発電所における実証試験
3.ユーグレナワックスエステルの特性
ユーグレナは、代替バイオ燃料のリソースとしても注目されている。好気培養条件下においてユーグレナは光合成によりパラミロンを大量に蓄積する。その量は最大乾燥重量当たり約50%に達する。好気培養したユーグレナを嫌気条件に移行することで、パラミロンは貯蔵脂質ワックスエステルへと変換される。ユーグレナのワックスエステルは、ミリスチン酸C14とミリスチルアルコールC14からなるミリスチルミリステートC28を主成分としており、他の微細藻類や油糧植物の生産する主要な脂肪酸は鎖長C16やC18であることから、C14を大量に生産可能な点は非常にユニークである。ミリスチン酸はディーゼルエンジンの耐ノック指数のセタン価が66.2と比較的高いため、一般的なバイオディーゼル燃料としての利用が可能な上、C14程度の軽質な脂肪酸はジェット燃料としての利用も期待できる。実際にユーグレナ油脂を水素化・異性化精製した試製油は、合成ジェット燃料規格 (ASTM D7566) に適合しており、ドロップインフュエルとして既存のジェット燃料と混合利用可能であることが確認されている。ワックスエステルの熱量は38.3 MJ/kgであり、エタノール (26.8 MJ/kg) と比較してもエネルギー密度の高い燃料を製造できること、またトリアシルグリセリドと異なり燃料精製段階でグリセロールなどの副産物が出ないことも利点である。
パラミロンからワックスエステルへの転化には、‘ワックスエステル発酵’と呼ばれる他に例を見ない独自の代謝系が機能している。ユーグレナを好気培養から嫌気条件に移すことで、パラミロンはグルコース単位に分解された後、解糖系を経てピルビン酸に変換され、ミトコンドリアに存在する脂肪酸合成系により脂肪酸を生成し、最終的にミクロソームにおいてワックスエステルが合成される。ミトコンドリアにおける脂肪酸合成経路は通常の脂肪酸合成経路とは異なり、マロニルCoAを用いずアセチルCoAをC2供与体とするβ-酸化系の逆反応に相当するが、β-酸化で不可逆反応に働くアシルCoAデヒドロゲナーゼに代わってtrans-エノイルCoAレダクターゼ (TER) が機能している。最近著者らのグループは次世代シーケンサーによるユーグレナ発現遺伝子情報を解析し、約157,000の転写物情報からこれまで未同定であったパラミロン合成酵素やTERを含めた脂肪酸代謝経路の構成酵素遺伝子を得ている (未発表)。現在これら酵素遺伝子の機能解析が進行中であり、好気/嫌気条件に応答したパラミロンとワックスエステルの代謝調節機能の解明が期待される。
4.形質転換によるパラミロン・ワックスエステル生産効率化の可能性について
上記のようにワックスエステル発酵はユーグレナに特異的な代謝系であり、ワックスエステルの燃料としての有用性から、今後のユーグレナを利用した産業として燃料生産が大きな期待を受けている。燃料生産のためのユーグレナの生産効率を高めるには、ハードウェアの設計と培養設備のコントロールによるパラミロン生成・蓄積条件およびワックスエステル発酵条件の最適化などが増殖効率の向上と共に重要となる。また、ワックスエステル合成のための代謝経路が遺伝子レベルで明らかになってきたことから、代謝工学に基づくパラミロンやワックスエステル生産の高効率化も重要な選択肢のひとつとなる。
昨今の遺伝子工学技術の発展に伴い、ユーグレナにおいても外来遺伝子の導入による形質転換技術が確立しつつあり、我々は代謝工学的手法によるバイオマス増産も試みている。その一例として、ユーグレナ細胞に対して藍藻FBP/SBPase遺伝子の導入が試みられた。FBP/SBPaseはカルビン回路の構成分子であるフルクトース-1,6-ビスフォスファターゼ (FBPase) とセドヘプチュロース-1,7-ビスフォスファターゼ (SBPase) が融合した藍藻で発見された新規酵素であり8)、植物への導入により光合成効率が顕著に改善することが報告されている9)。藍藻FBP/SBPase遺伝子のユーグレナへの導入により、野生株に比べ光合成活性の上昇とそれに伴うパラミロンやバイオマス量の増加が認められ、その有用性を実証している (投稿準備中)。一方、ユーグレナ由来のワックスエステル合成酵素を導入した酵母にワックスを生産させた例も存在しており10)、ワックスエステル合成はユーグレナに特有な代謝形態ではあるものの、他の微生物の代謝経路との一部互換性があることも示唆される。このため、セルフクローニングによるワックスエステル合成経路の強化以外にも、他の微生物由来の外来遺伝子を積極的に導入することで燃料生産の効率を向上させることも期待できる。
5.今後の課題
微細藻類による物質生産については古くから研究が行われており、将来性も期待されるが、産業のスケールでの燃料生産には至っていないのが現状である。藻類バイオマスからの燃料生産を産業とするためには、ユーグレナを含めた藻類の安定的かつ大規模に安価に生産する技術の確立が求められる。そのための一つの手段として、代謝工学の技術が環境変化に対するロバスト性の付与や目的油脂の高効率な産生に大きく寄与するはずである。また、従来の原油の精製においてもナフサからガソリン、ジェット燃料、ディーゼル燃料、重油、コールタールなどを供給してすべてを有効に販売して現在の流通するコストを実現しており、藻類由来の燃料生産においても燃料以外の高付加価値の副産物を利用することでコスト競争力を持たせることが重要と考えられる。藻類による燃料生産を軸にした炭素循環型社会の構築は持続社会構築の大きな一翼を担うと考えているので、筆者らのグループに限らない今後の継続的な技術開発と社会への応用に期待をしたい。
文献
1) 山田常雄ほか編:岩波生物学辞典(第3版), 岩波書店 (1983).
2) 中野長久, 浜崎和恵, 竹中重雄, 宮武和孝, 谷 晃, 相賀一郎:CELSS学会誌, 7, 15(1995).
3) Clarke, A. E., Stone, B. A.: Biochim. Biophys. Acta, 44, 161(1960).
4) Inui, H., Miyatake, K., Nakano, Y., Kitaoka, S.: FEBS Lett., 150, 89(1982).
5) 朝山雄太, 鈴木健吾: FOOD Style 21, 17, 73(2013).
6) 近藤次郎: 環境科学会誌, 26, 417(1996).
7) 鈴木健吾, 尾形直久: Biophilia, 3, 56(2007).
8) Tamoi, M., Murakami, A., Takeda, T., Shigeoka, S.: Biochem. Biophys. Acta, 1383, 232(1998).
9) Miyagawa, Y., Tamoi, M., Shigeoka, S.: Nat. Biotechnol., 19, 965 (2001).
10) Teerawanichpan, P., Qiu, X.: Lipids, 45, 263 (2010).
![]()