�y�g�s�b�N�X�z
��K�͐A���H��ɂ�����c�I�ʃ��{�b�g�V�X�e���ƊT�����Y���f�f
���c�O�a�A��s����A�L������
��{��@�E�H�A��{��@�E�H�A��{��@�E�H
�P�D�͂��߂�
���E���ňُ�C�ۂ�������ɂ��y��̍r�p���N�X�[�������A�_�Ɛ��Y�̒���뜜����Ă���B���̂悤�ȏ�Ŕj����V�����_�Ƃ̎�i�Ƃ��ĐA���H�ꂪ�ߔN���ڂ���Ă���A����Ƃ��Č����E�J�����i�߂��Ă���1)�B���ɋߔN�J�����ڊo�܂����̂��A�l�H���^�A���H��ł���B�l�H���^�A���H��́A���E���x���x�E��_���Y�f�Z�x�E�|�{�t�g���Ȃǂ͔̍|�ɕK�v�Ȋ�������S�Ď{�ݓ��Ő���ł��A�G�߂�ꏊ�ɑ���ꂸ�ɐA���̈��萶�Y�������ł��闘�_������1)�B���̗��_�ɂ��A�����n�т⊦��n�ȂNJC�O�̍k��s�K�n�ł̐A�����Y���\�ƂȂ邽�߁A�A���H��͗A�o�Y�ƂƂ��Ă����҂���Ă���B�������Ȃ���A�l�H���^�A���H��͏Ɩ��ƋǗ��ɖc��ȓd�͂�K�v�Ƃ��邽�߁A�����j���O�R�X�g�̒ጸ���ő�̉ۑ�ƂȂ��Ă���B���̑�Ƃ��āA����d�͂�70%���߂�����ɑ��ďȃG�l����}�邱�Ƃ�A�������ɂ��ȗ͉����������邪�A���Y�V�X�e���̑�K�͉��ɂ�鐶�Y�H���̍����������d�v�ȋZ�p�ۑ�ƂȂ��Ă���1)�B
��K�͐A���H��ł́A��q�E�c���̃T���v���W�{�����c��ƂȂ�A���̌��ʁA�c�̌̍��͈��肵�����v�ʂƂ��Č���Ă���B���̌̍������ƂɗD�ǂȕc��I������̂��c�I���ł���A���������ɂ����ėL���Ȏ�i�ƂȂ�B�܂��A��K�͐A���H��ł͖�������{�ɋy�ԕc�I�ʂ��K�v�ł���A���̏����ʂ́A�p�����ƐM�����̖ʂ���l��ɂ���Ƃ͍���ł���@�B�����K�{�ƂȂ�B
��ʂɑ�K�͂ȕc�f�f�ɂ́A�ėp�������������������\�ȉ摜�v���@���̗p����Ă���B�ʏ�A�摜�ɂ��f�f�́A�c�̌`���傫�����w�W�Ƃ��B���͈�x�ł���B�������A�f�f���x�̌���̂��߂ɂ́A������v�� (���n��v��) ���L���ł��邱�Ƃ����X�ɂ��Ă���B���̗L�����́A�T�����Y���ƌĂ����������̐��̃��Y���������̐��̏��Ɋ܂܂�邱�ƂɗR������2)�B�Ⴆ�Ό��������x������x�́A���E���x�����̍P��I�Ȋ����ɂ����Ă��A����������Ƃ��đ������J��Ԃ����Ƃ��m���Ă���B�]���āA�T�����Y�����炻�̐U��������Ȃǂ̓����ʂ𒊏o���A�c�f�f�̎w�W�Ƃ��邱�Ƃ��f�f���x����ɂ����ėL���ł���3)�B
����ŁA�A���H��̑�K�͉��ɔ����A�K�R�I�ɍ͔|���͍L���Ȃ�A���̐���͂܂��܂�����Ȃ�B���ɁA���x���z�̕s�ψꉻ�⎞�ԕϓ��Ȃǂ��������郊�X�N�����܂�1)�B�A���̐����́A��������܂߁A���x�ɑ��Ċ��������A�A���̊T�����Y�������x�ɑ��Ċ����������Ƃ��m���Ă���4)�B���������āA���x�ϓ����T�����Y���ɋy�ڂ��e����]�����邱�Ƃ́A�c�I���̐��x����ɂ����ďd�v�ȉۑ�ł���B
�{�e�ł́A��K�͐A���H��ɂ�����ŐV�̕c�I�ʃ��{�b�g�V�X�e�����Љ��B���̃V�X�e���̓����́A����5,000���̕c��ΏۂɎ��n��I�Ȑf�f���s���A�T�����Y���̐f�f�����{�ł���_�ł���B�܂��A��q�̂悤�ɊT�����Y���͉��x�ɑ���q�����������߁A�c�f�f���ɂ����č����x�ȉ��x���߂��K�v�ƂȂ�B�ŋߓ��������Ŗ��炩�ɂ������x��炬���T�����Y���ɗ^����e���ɂ��Ă��Љ�����B
�Q�D��K�͐A���H��ƕc�I�ʃ��{�b�g�V�X�e��
�}1�́A�o�ώY�Əȁu�C�m�x�[�V�������_���n�������Ɓv�̕⏕���A2014�N9���ɏv�H�������{����w�̑�K�͐A���H��ł���B���Y5,000���͔̍|�\�͂������������ł��L���̑�K�͐A���H��ł���B���{�݂ł́A�����j���O�R�X�g40�����̎�����ڕW�Ƃ��A����3�̎�Ȏ�g���s���Ă���B�@�c�I�ʃ��{�b�g�V�X�e�� (���������j�A�A�������͔|�V�X�e�� (�ȗ͉�)�A�B�͔|�����̊��SLED�� (�ȃG�l��) �ł���B�@�A�͓��ɍ���J�����ꂽ�Ǝ��̃V�X�e���ƂȂ��Ă���1)�B
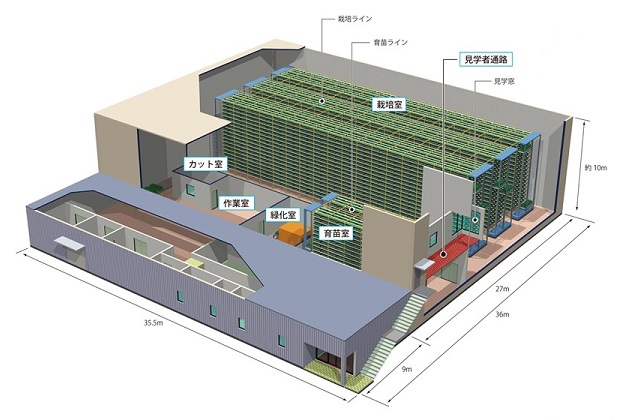
�}1�@���{����w�@�V����A���H��1)
�����ł͕c�I�ʃ��{�b�g�V�X�e�����Љ��B�I�ʕꐔ��6,000�{�^���Ƃ��A�D�G�c�Ƃ���5,000�{�^����I�ʂ���B�c�I�ʂ́A�Ή����ɂ�����4���Ԃ̔d��E�Ή��H���̍ŏI�i�K�ōs���A�d��A��o���A�Ή��A�c�I�ʂ̏��ōs���B����6,000�{�Ƃ��������ʂ́A�p�����ƐM�����̖ʂ���@�B�����K�{�ƂȂ�B
�ȉ��͐f�f�X�L�[���̊T�v�ł���B�d��E�Ή��H���ł́A������2���ԈÎ��ɂĉ�o�����s���A���̌�4���Ԕ��FLED���ɂ��Ή����s���B�Ή��̍ŏI�� (�Ή��H����4���ڊJ�n����4���ڏI����) �ɂ����āA�Ή��Ɠ����i�s�ŕc�f�f���s���B�ŏI����6,000�{�̕c�̌������s���A�c�f�f�ŏ�ʂɃ����N������̂���5,000�{��I������B�d��͐�p�̔���p�p�l�� (300�{�^�p�l���A�p�l���T�C�Y600�~300 mm�A�|�n�F�E���^���}�b�g) �ɂčs���A�Ή��̏ȃX�y�[�X���ƍ�Ƃ̌�������}��B�S�Ă̔����ߒ��ɂ����āA2�̃p�l����1�P�ʂƂ����戵�����s���B�Ή����A�����ĕc�f�f�V�X�e���ւ̈ړ��́A�S�Ď����������u��p������̃v���O�����ɏ]���čs����B
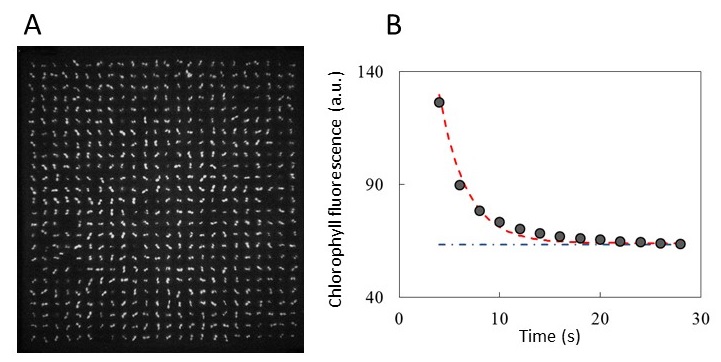
�c�f�f�̃V�X�e���́A�����x�E���𑜓x�̗�pCCD�J�����ƃN�����t�B����N�p�̐FLED���u����\�������B����p�p�l�����ƈÔ����Ɏ����������u�ňړ������A�FLED�����N���Ƃ��āA�A���̂̃N�����t�B���F�f���N������B�N�����t�B�����琶����u����4���Ԗ���1����6��B�e���� (�}2)�B�v��1����5���Ԃ̓���f�[�^���擾���A�̃T�C�Y�A�`��A�N�����t�B���u�����v������B���̂悤�Ɏ擾���ꂽ1��6�_�̎��n��f�[�^����A�@�̃T�C�Y�A�A�`����Ă̌`�ԃf�[�^�A�B�N�����t�B���u�����x�A�C�T�����Y���̐U���A�D�T�����Y���̎����Ȃǂ��Z�o����B�@�`�D��c�f�f�Ɋւ����b�f�[�^�Ƃ��A����̕]������p���ĕc�̗D�ǐ��𐔒l������B�c�f�f�̕]�����́A�@�B�w�K�Ȃǂ����p���Ē�߂�B����6,000�{�̕c�f�f���s���ƖN��106�̂̃f�[�^��~�ςł��A���̖c��ȕc�f�f�f�[�^�́A��w�������̃T�[�o�[�ŕۊǁE��͂���A�c�f�f�̃p�����[�^�X�V��V�K�A���S���Y���J���ɗ��p�����B

��q�̐f�f��Ƃ�24���ԘA�����čs���A�I�ʒ��O�ɑS�̂ɗD�ǐ��̐��l���t�^�����B�S�̂�ID�ƕt�^���ꂽ�D�ǐ��̐��l��������A���{�b�g�ɓ`�����A���5,000���̑I�����s���A��c�p�̃p�l���ɒ�A����B������A���{�b�g�́A��{�I�ɂ̓}�j���A������ɂ���ċN��������̂Ƃ��A��Ǝ҂̓����̂��ƂŃp�l���ꖇ���ƂɎ�����A���s���B��Ǝ҂́A��A�����̃p�l�������X�e�[�W�̈�c���ֈړ���������A���̎����I���E��A���s���B��Ǝғ����Ƃ��邱�ƂŁA���G�Ȕ����@�\���ȗ��ł��A�����V�X�e���̊J���E����ɂ�����ߏ�ȃR�X�g��}���邱�Ƃ��ł���B�Ȃ��A��c�p�p�l���́A��c�̏ȃX�y�[�X���ƍ�Ƃ̌��������l�����A150�c�^�p�l���A�p�l���T�C�Y890×590 mm�ƂȂ�B
�c�f�f�ɂ����ĊT�����Y���ɒ��ڂ��Ă���_�͐��E�I�ɂ����߂Ă̎��݂ł���A����ɂ��c�f�f�̐��x�̌����ڎw���Ă���B
�R�D������ɂ�����T�����Y��
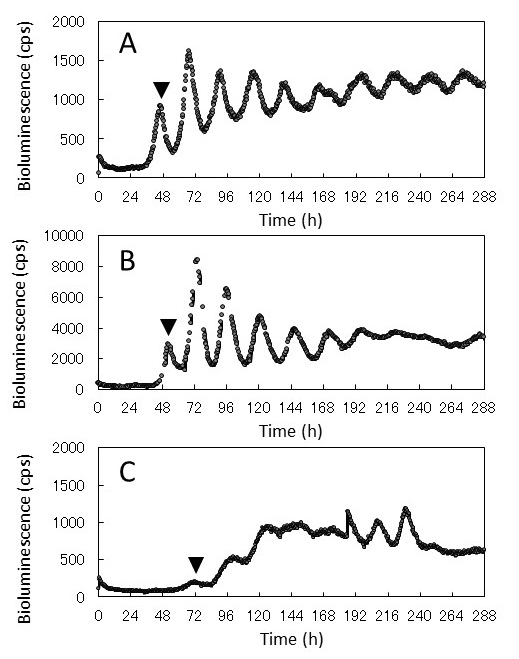
�M�҂̃O���[�v�ł́A�A���H��̋Z�p�J���Ɠ����ɁA�A���̊T�����Y���ɂ��Ă�������i�߂Ă���B�}3�́A��`�q�g�����O���[���E�F�[�u���^�X (AtCCA1::LUC) ��p���Čv������������ɂ�����T�����Y���̗l�q�ł���5-7)�B�����ł̓��V�t�F���[�[��`�q�����|�[�^�Ƃ��ėp���A��`�q�̔����ʂ��j��Ōv�����Ă���B�v�����u�ɂ�Kondotron�ƌĂ��n�C�X���[�v�b�g�������������v�����u��p�����B���V�t�F���[�[�����̎��n��f�[�^�͐A���̂ɂ���ĈقȂ���������B�}3A�̌̂ł́A�����悻t=48 h�Ń��V�t�F���[�[�����ʂ��}�����A���U���������Ă���Bt=48 h�ɂ����锭������́A�������m�F����鎞�Ԃƈ�v����B����A�}3B�̌̂ł́A���U������������At=216 h�ȍ~�ɂ����ĐU�����������������Ă���B�܂��}3C�̌̂ł́A���U�����������A�U�����r���ő傫���Ȃ�Ȃ��}3AB�Ƃ͈قȂ锭�����������B�{�����ł͍��v80�̂̔����v���������s�������A�}3A�Ɠ��l�̔����p�^�[�����������̂�37�́A�}3B��25�́A�}3C��13�́A���肵�Ȃ������̂�5�̂ł������B

�S�D�W�c�����Ɖ��x��炬
��q�̂悤�ɁA�T�����Y���͉��x�ϓ��ɑ��Ċ������B���̂��ߍ͔|���ɂ����ĉ��x���ϓ�����ƊT�����Y���v���ɂ����ăm�C�Y������B�ȉ��ł́A�X�̕c�����T�����Y���̏W�c�����ɒ��ڂ��āA���x��炬���T�����Y���v���ɂ�����m�C�Y�ƂȂ邱�Ƃ��Љ��B
�}4�́A�͔|���x�̗�ł���B�}4A�͉��x�����ɐ��䂵���ꍇ (���䉷�x���� (a))�A�}4B�͉��x������s���Ă��Ȃ��ꍇ (����s�����x���� (b)) �ł���B�܂��A�T�����Y���̏W�c������R�����̂悤�ɒ�`���ĕ\�����Ă���9)�B
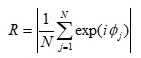
������øj�͕cj�̊T�����Y���̈ʑ��AN�͕c�̑����ł���B���䉷�x���� (a) �ɂ����鉷�xTa��Ta=26�}0.2���łقڈ��̉��x���ۂ���Ă��� (�}4A)�B���̏����̉��œ�����Ra�́A���菉�� (t=48 h�t��) �ɂ�����Ra=0.7�t�߂̔�r�I�����l�������Ă��邪�A���̌㌸�����Ă����ARa=0.2�`0.5�𐄈ڂ����B�}4A����A���������Ɠ����ȏ�̓������ɉł��Ȃ����Ƃ�������B����́A�̊Ԃ̓����𑣂��v�������݂��Ă��Ȃ����Ƃ��Ӗ����Ă���B
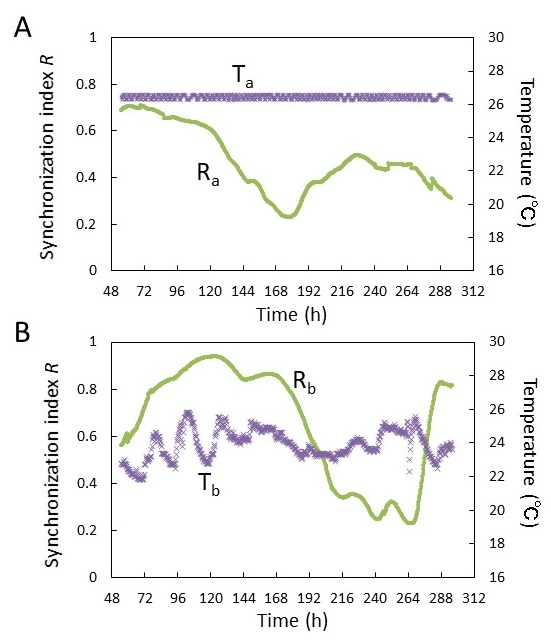

����A����s�����x���� (b) �ő��肳�ꂽ���xTb��Tb=23.8�}2.0���ŕϓ����傫�����Ƃ������� (�}4B)�B����t=72�`144 h�ɂ����ē������̉��x�ϓ� (�ő剷�x��3��) ��At=264�`288 h�ɋ}���ȉ��x�ቺ (���x��3��) ��������B������Rb�͔��菉�� (t=48 h�t��) �ɂ�����Rb=0.6�������Ă���B����́A���䉷�x���� (a) �ɂ����铯����Ra�Ɠ����x�̒l�ł���B���̌�At=72�`144 h�̎����I���x�ϓ����݂��鎞���ł�Rb=0.94�܂ŏ㏸���A�܂�t=264�`288 h �ɂ����鉷�x�ቺ���ɂ����Ă��A�}���ȓ������̏㏸���݂��A���̌㍂��������Rb=0.83�������Ă���B
�ȏ�̂��Ƃ���A���x�ϓ����Ȃ��ꍇ�A�������͎��ԂƂƂ��ɒቺ���邪�A���x�ϓ�������ƒ����ɏW�c�̓��������������邱�Ƃ�������B
�T�D�A���H��ɂ����鉷�x�Ǘ��ƊT�����Y��
�A���H��ɂ���������I�ȉ��x�ϓ����A���͔|�ɗ^����e���ɂ��ďq�ׂ�B�c�I���ɂ����ẮA�A���̊T�����Y�����������ƂɌ̊Ԃ̏����I�Ȑ����̍��ق肵�Ă���B�������Ȃ���A�͔|���Ɏ����I�ȉ��x�ϓ���������ƌ̓��m�����x�ϓ��ɓ������Ă��܂��A�e�A���̂����T�����Y�������̍�����������B���̌��ʁA�T�����Y�������ɂ��̍��̔��ʂ�����ɂȂ邽�߁A�c�I�����s����c�i�K�ł́A���x�ϓ��̔������ł��邾���}����悤���x�Ǘ������Ȃ���Ȃ�Ȃ��B
�܂��t�ɁA�͔|���ɂ����ẮA���x�ϓ����̊Ԃ̓����𑣐i���邽�߁A�͔|�A���̊T�����Y����Ĉ�I�ɐ��䂷��ۂɑ傫�ȗ��_�ƂȂ�B���̈���ŁA��K�͂ȍ͔|���ł݂͌��ɗ��ꂽ�͔|��̊Ԃɉ��x�ϓ��̈ʑ������\��₷���B�T�����Y���̐���͌��ɂ���Ă��s���邽�߁A���̖��Â̈ʑ��Ɖ��x�ϓ��̈ʑ��̊Ԃɂ����������邱�Ƃ��l������B���̂悤�ȏꍇ�A�͔|���S�̂ɓn��A���̊T�����Y�����ψ�ɐ��䂷���Ƃ��ł����A�A���̐����̓x�����A���Ȃ킿�i���̋ψꐫ�ɉe�����o�Ă��܂��\��������B�ȏ�̗��R����A�c�I���ɂ����Ă͉��x�ϓ��̔������ł��邾���}���A�܂��͔|���ɂ����Ă͉��x�s�ψꐫ�������Ȃ��悤�A���x�̍������x�Ǘ������߂���B
�U�D������
�����̓V��s�ǂ͍앨�̍����������N�����A���ʓI�ɐl�H���^�A���H��ւ̎��v����ɂȂ����Ă���B�A���H��̋Z�p�J���͂������N�ŋ}���ɐi�W���Ă��邪�A�܂��܂��ۑ�͑����B�R�X�g�ጸ�̖ʂ����K�͉����K�{�ł���A����ɔ�����Ƃ̎���������ψꉻ�̋Z�p�J�����K�v�ł���B����܂ł��A�͔|�������Z�p�������Z�p�ɂ��Ă̌����J���͍s���Ă������A�\���Ȑ����f�[�^�Ɋ�Â������J���ł������Ƃ͌�����B�e��Z�p�J���́A�����܂őΏۂƂȂ�A���̏�Ԃ��k���Ɍv�����͂��A�œK���Z�p��v���ׂ��ł���B�{�e�ŏЉ���T�����Y���̉��x���ɒ��ڂ�������w�j�́A���̈��ł���B
�ߔN�A�ԗ��I�Ȉ�`�q�������܂ޖc��Ȑ����f�[�^�̎擾��~�ρA�����Ă��̉�͂ɕK�v�Ȏ��Ԃ�R�X�g�͌��I�ɏ������Ȃ��Ă��Ă���B����A��K�͐����f�[�^�Ɋ�Â��œK���Z�p�̐v�́A�\���ɒ���ł���ƂȂ��Ă���ƌ�����B�������ӂƂ��c��Ȑ����f�[�^��e�ՂɎ擾�ł���A���H��Ƒ�K�͐����f�[�^�����̗Z���́A�V���Ȋw�p�Ƃ��Ė��͓I�ł��邾���łȂ��A�V���ȎY�ƋZ�p�̊�b�Ƃ��Ċ��҂����B
�ӎ�
�{�����́A�o�ώY�ƏȃC�m�x�[�V�������_���n���Ƃ�JST A-STEP���݉��A�����Ȋw�ȉȊw������⏕�� (No. 25712029, No. 25119721) �ɂ��x�����čs�����B
����
1) Takatsuji, M., Kozai, T. (editorial supervision): �hImportant Task and Countermeasure of Plant Factory Management�h, Institute for Information. (2014), (in Japanese).
2) Farre, E. M.: Plant Biol., 14, 401 (2012).
3) Fukuda, H., Ichino, T., Kondo, T. Murase, H.: Environ. Control Biol., 49, 51 (2011).
4) Thines, B., Harmon, G. F.: Proc. Natl. Acad. Sci. USA, 107, 3257 (2010).
5) Ukai, K., Inai, K., Nakamichi, N., Ashida, H., Yokota, A., Hendrawan, Y., Murase, H., Fukuda, H.: Environ. Control Biol., 50, 237 (2012).
6) Higashi, T., Kamitamari, A., Okamura, N., Ukai, K., Okamura, K., Tezuka, T., Fukuda, H.: Environ. Control Biol., 52, 21 (2014).
7) Udo, R., Moriyuki, S., Ukai, K., Fukuda, H.: Trans. of the JSRAE, 31, 123 (2014), (in Japanese).
8) Fukuda, H., Murase, H., Tokuda, I.: Scientific Reports, 3, 1533 (2013).
![]() �@
�@